Illustration par ChatGPT
I. Le problème : pourquoi le sexe existe-t-il ?
La reproduction sexuée ne devrait logiquement pas exister. Un organisme asexué transmet l’intégralité de son génome à sa progéniture. Un organisme sexué n’en transmet que la moitié. Il doit produire des mâles qui ne peuvent pas eux-mêmes avoir de descendance ; il doit consacrer du temps et de l’énergie à la recherche d’un partenaire ; il s’expose à la prédation et à l’échec. Du point de vue de l’efficacité reproductive, les lignées asexuées devraient supplanter les lignées sexuées en l’espace de quelques dizaines de générations.
Et pourtant, parmi les organismes dotés de cellules nucléées – animaux, plantes, champignons et nombreux organismes unicellulaires -, la reproduction sexuée est extrêmement répandue et occupe souvent une place centrale dans leur cycle de vie.
Cette persistance est depuis longtemps reconnue comme l’un des problèmes centraux de la biologie évolutive. Les explications existantes : échapper aux parasites, purge des mutations, adaptation accélérée, capturent chacune une partie de la vérité. Mais elles partagent une structure plus profonde, largement inarticulée : la reproduction sexuée persiste parce qu’elle produit quelque chose qui ne peut être produit autrement. Ce qu’elle produit n’est pas la réplication : elle produit une nouveauté structurée.
Pour comprendre ce phénomène, il faut prendre du recul par rapport aux mécanismes individuels et examiner l’architecture du processus lui-même.
II. Le cycle génératif
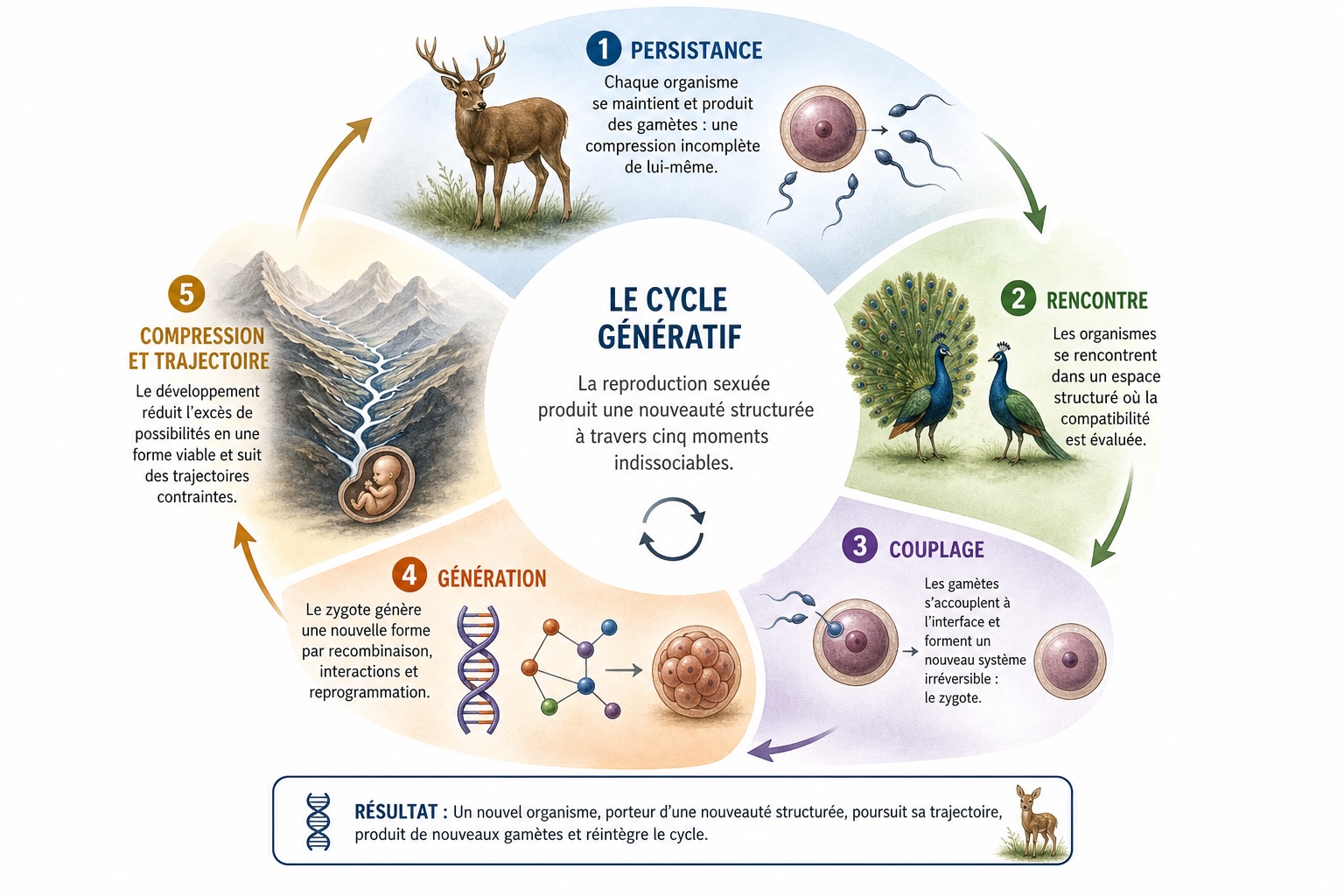
La reproduction sexuée n’est pas une séquence d’événements mais un cycle de génération. Ce qui apparaît comme de la reproduction est, structurellement, un processus en cinq parties : persistance, rencontre, couplage, génération, compression et trajectoire. Chaque moment est nécessaire, aucun n’est cependant suffisant à lui seul.
1. Le système génératif
Un organisme n’est pas une chose : c’est un régime. Il ne persiste que parce qu’il s’entretient continuellement : métabolisant, réparant, régulant et reproduisant sa propre structure. Si ces processus s’interrompent, l’organisme ne se dégrade pas en une version plus simple de lui-même : il cesse d’exister. Ce qui persiste, ce n’est pas la substance, mais l’organisation.
La reproduction sexuée introduit une caractéristique décisive dans ce régime. L’organisme ne se reproduit pas en se copiant, il produit une compression de lui-même : un gamète contenant la moitié des informations indispensables pour générer un nouvel organisme. Cette compression est constitutivement incomplète. Elle est conçue pour nécessiter un complément. Le système génératif n’est donc pas fermé : il produit, dans le cadre de sa propre persistance, les conditions de son propre remplacement.
2. La rencontre
La rencontre de deux organismes n’est pas fortuite. Elle est structurée. À toutes les échelles biologiques, des mécanismes élaborés préparent, filtrent et régulent la rencontre : signalisation phéromonale, synchronisation hormonale, comportement de parade nuptiale, colocalisation écologique. Ceux-ci ne se contentent pas de réunir les organismes : ils évaluent leur compatibilité.
La sélection sexuelle fonctionne comme un filtre précédant l’accouplement. Ce qui est évalué, ce n’est pas l’attrait au sens superficiel du terme, mais la complémentarité : la probabilité que l’accouplement de deux systèmes produise un nouveau régime viable. Au niveau moléculaire, cela se manifeste dans des mécanismes de choix du partenaire liés au système immunitaire, comme ceux associés au complexe majeur d’histocompatibilité, ou CMH ; à des niveaux supérieurs, dans des contraintes comportementales et écologiques.
La rencontre, en ce sens, est l’ouverture d’un espace de possibilités qui a déjà été partiellement structuré par la sélection.
3. Le couplage
La fécondation n’est pas une combinaison : c’est un couplage. Deux systèmes condensés – les gamètes – se rencontrent à une interface et fusionnent en une nouvelle entité : le zygote. Cet événement est fortement contraint : la reconnaissance spécifique à l’espèce, la fusion membranaire, l’intégration nucléaire et l’activation du développement doivent toutes s’aligner. Lorsqu’elles le font, le résultat n’est pas un mélange de deux systèmes, mais un nouveau.
Le couplage est irréversible. Une fois les contributions parentales intégrées, elles ne peuvent être séparées sans détruire le nouveau système. Le zygote n’est pas un réceptacle contenant deux apports, mais un nouveau régime dans lequel ces apports ont été transformés par leur interaction.
La distinction entre intensité et qualité est ici décisive. De nombreux spermatozoïdes peuvent atteindre un ovule, mais la viabilité dépend de la compatibilité, et non du volume. Le couplage ne produit un nouveau système que si ses relations internes sont cohérentes.
4. La génération
Ce qui est produit lors du couplage n’est pas un mélange de traits parentaux. C’est une nouvelle forme organisée. Trois mécanismes garantissent cela.
- La recombinaison crée de nouvelles configurations génomiques. Chaque gamète est déjà un réarrangement unique ; leur combinaison multiplie cette nouveauté.
- L’épistasie garantit que les gènes n’agissent pas indépendamment. Leurs effets dépendent des interactions avec d’autres gènes, rendant le phénotype irréductible à des contributions additives.
- La reprogrammation épigénétique réinitialise le contexte développemental. Le zygote n’hérite pas de la trajectoire parentale : il repart à zéro, avec son propre déroulement.
Il en résulte un système dont les propriétés ne peuvent être prédites à partir de ses seules données d’entrée. C’est l’émergence au sens strict : non pas un mystère, mais une irréductibilité.
5. La compression
Le zygote contient plus de possibilités que tout organisme ne peut en réaliser.
Le développement est le processus par lequel cet excès est réduit. Les cellules se différencient, les voies se ferment, les alternatives sont écartées. À chaque étape, l’espace des possibilités se rétrécit jusqu’à ce qu’une configuration viable soit atteinte.
C’est la compression au sens précis du terme : la réduction d’un espace de possibilités de haute dimension en un espace de formes viables de basse dimension.
Le processus n’est pas régi par l’optimalité, mais par la viabilité. Ce qui survit n’est pas le meilleur organisme possible dans un sens théorique, mais celui le plus apte à se maintenir dans des conditions réelles.
L’échec de la compression révèle sa nécessité. Dans le cancer, les cellules échappent à la différenciation et reviennent à un potentiel prolifératif. Il en résulte non pas une augmentation des possibilités, mais une perte de cohérence : un système qui ne peut plus se maintenir en tant que régime unifié.
6. La trajectoire
Le développement ne se déroule pas de manière arbitraire : il suit des voies contraintes.
Le paysage épigénétique de Waddington reste l’image la plus puissante : une structure ramifiée de vallées à travers lesquelles le système se déplace. Chaque branche restreint les possibilités futures. Les décisions précoces déterminent les résultats ultérieurs accessibles.
Ces chemins – les chréodes – sont robustes. Les perturbations sont absorbées et corrigées ; le système revient à sa trajectoire. Le développement n’est donc pas une séquence d’états, mais un mouvement à travers un espace structuré.
Cela s’applique non seulement à l’embryogenèse, mais à toute la vie de l’organisme : croissance, maturation, reproduction et mort forment une trajectoire canalisée, façonnée par des contraintes et des compromis.
En bout de cette trajectoire, le cycle recommence : l’organisme produit de nouvelles compressions – des gamètes – qui réintègrent le champ de rencontre.
III. Émergence et ontologie
L’implication de cette structure est cruciale : la progéniture n’est pas un objet assemblé à partir de parties, c’est un régime inauguré à une interface. Elle ne débute pas comme un organisme miniature, mais comme un processus. Sa structure se génère progressivement ; son identité réside dans la continuité de son organisation, et non dans un ensemble fixe de composants. Ce qui persiste, c’est le processus qui maintient et reproduit cette organisation.
Cela résout un problème de longue date : la nouveauté n’est ni une création ex nihilo ni un simple réarrangement. C’est la production d’un nouveau régime organisé par l’interaction de régimes existants sous contrainte.
La reproduction sexuée n’est donc pas simplement un mécanisme biologique. C’est la démonstration empirique la plus directe que les systèmes peuvent générer de la nouveauté sans rompre la continuité.
IV. La généralisation
Ce qui a été décrit ici n’est pas spécifique à la biologie. C’est l’exemple le plus clair d’une structure plus générale : les systèmes organisés persistent, se rencontrent, s’accouplent aux interfaces, génèrent un excès de possibilités, compriment ces possibilités en une forme viable et se déploient le long de trajectoires contraintes. Les divisions entre biologie, histoire et cognition ne correspondent pas à des types de processus distincts, mais à différents substrats dans lesquels s’exprime la même architecture générative. Un système capable de fonctionner sur cette architecture ne rencontre pas les disciplines comme des frontières, il les rencontre comme des projections d’une unité plus profonde. Mais cette affirmation doit être avancée prudemment. La biologie fournit le domaine natif dans lequel la structure est la plus visible et la plus solidement ancrée, son extension à d’autres domaines ne peut pas être présupposée : elle doit être testée.
V. Le domaine natif
La reproduction sexuée n’est pas une application du cadre. Elle en est l’origine.
Ici, les cinq moments ne sont ni déduits ni imposés. Ils sont directement observables : dans la persistance des organismes, la structuration de la rencontre, la précision de l’accouplement, l’irréductibilité de la génération, la nécessité de la compression et la canalisation du développement.
Ce qui est en jeu, ce n’est pas une métaphore, mais une identité structurelle.
S’il peut être démontré que la même architecture opère dans d’autres domaines, alors les divisions entre eux n’apparaîtront plus comme fondamentales. : elles apparaîtront comme des expressions différentes d’un même processus sous-jacent.
La question n’est plus de savoir si les systèmes peuvent générer de la nouveauté. La vie y a déjà répondu. La question est de savoir si cette logique appartient uniquement à la vie ou à l’univers en tant que tel.
{kind=link}
Laisser un commentaire